Como bala de cañón
Compartir
El tren imaginario en el que vamos se ha detenido. Como para tomar impulso, podríamos decir. Estamos a punto de internarnos en el núcleo de una célula eucariota. Finalmente. La última parada en el camino de ida de este viaje por el micromundo vegetal. O micromundos, mejor dicho, en plural. Un viaje por los escenarios de ese cosmos microscópico de las plantas, transitando por los senderos por donde avanza la ciencia en la comprensión de los mecanismos que intervienen en el desarrollo de la vida vegetal con el propósito de potenciar el rendimiento productivo y la adaptabilidad ambiental de las especies que cultivamos.
El desafío consiste en lograr mejores resultados prácticos, con mayor precisión, en menos tiempo y con efectos lo más duraderos posible. Un intento de domesticación de esas especies vegetales con las que nos alimentamos, que está en el origen mismo de la agricultura y que al cabo de miles de años estaríamos más cerca de poder inducir de manera inteligente, amigable, y en definitiva más eficaz. Y en especial -aunque falta todavía- por el mérito de haber avanzado en la comprensión de lo esencial: el lenguaje molecular de los organismos vivos.
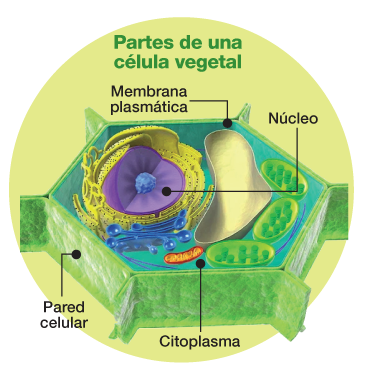
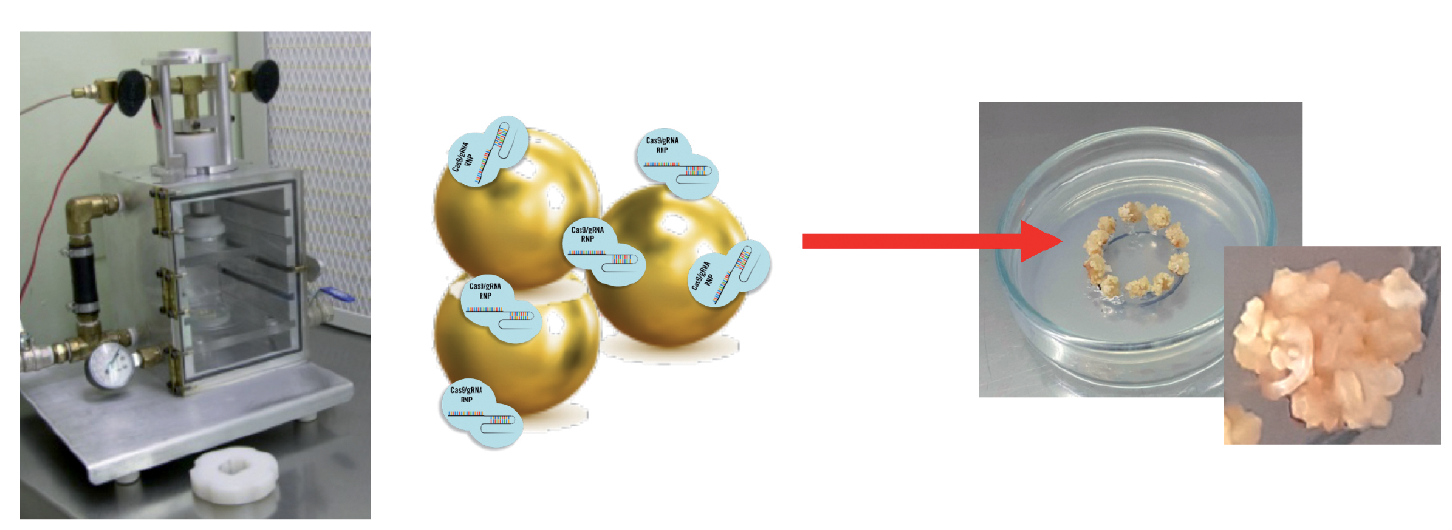
Estamos ahora a instantes de ser propulsados al interior del núcleo de una célula de caña de azúcar. Lo haremos montados en un chorro de alta presión de partículas de oro, revestidas con el material genético de diseño con las que se la impactará con la intención de modificar puntualmente su genoma. Si se logra (porque habrá obstáculos para sortear), se habrá conseguido introducir un cambio en el alfabeto de su ADN. Esa modificación le permitirá a la planta que de ahí resulte adquirir un nuevo atributo de su personalidad. Estamos a punto, esto es, de participar en una experiencia de edición genómica. Hasta aquí -y gracias a lo que aprendimos del sistema de defensa de las bacterias– el método más preciso de intervención en el ADN de las plantas.
Imaginamos la escena; imaginemos la exaltación que genera esa expectativa. Esta pausa previa, como para tomar aliento, nos da tiempo para pensar.
Las bases materiales del lenguaje celular
(Nos importa por lo que se sabe, pero también por lo que queda por saber)
Algo hemos aprendido en el camino hasta acá. Por ejemplo, que con el uso de cierta licencia y provisoriamente al menos, podemos hablar de algo así como de un lenguaje propio de las células vegetales. Usamos el término porque hoy ya sabemos que hay ahí algo que es, o se parece mucho a un lenguaje, y que entenderlo así ayuda en el camino de su desentrañamiento.
Hablamos (en lenguaje humano) del intercambio de señales bioquímicas que ocurren tanto dentro del citoplasma celular como entre el citoplasma y el núcleo, dentro del núcleo mismo y también entre células. Un sistema codificado de mensajes que transitan en un fluir constante de señales entre efectores y receptores celulares que reaccionan en consecuencia según su “significado”. Un sistema de mensajes cifrados en macro y micromoléculas compuestas a su vez por átomos de los mismos elementos químicos que constituyen la materia orgánica toda, cualquiera sea la función que a una determinada combinatoria le toque cumplir.
Esa función dependerá de qué átomos se combinan con qué otros formando esas moléculas que, además de estar “escritas” con esas “letras”, ocupan como estructura un lugar en el espacio celular. Su función dependerá en definitiva de las propiedades intrínsecas de su composición y de su arquitectura espacial, como si se tratara de ideogramas bioquímicos tridimensionales, que “hablan” también a través de su forma.
Ahí estarán, como principales protagonistas, el carbono (C), el hidrógeno (H), el oxígeno (O), el nitrógeno (N), el azufre (S), el fósforo (P), en distintas combinaciones, aunque no siempre todos con todos, y asociados con otros, orgánicos y minerales, formando parte de la contextura de la planta, de sus hojas, de sus vasos, y también de las estructuras moleculares que cumplen especialmente funciones de memoria informativa y comunicación en un sistema que, como dijimos, bien puede concebirse como un lenguaje. Ese listado de elementos no es infinito, pero la magnitud de sus posibilidades expresivas es todavía incalculable.
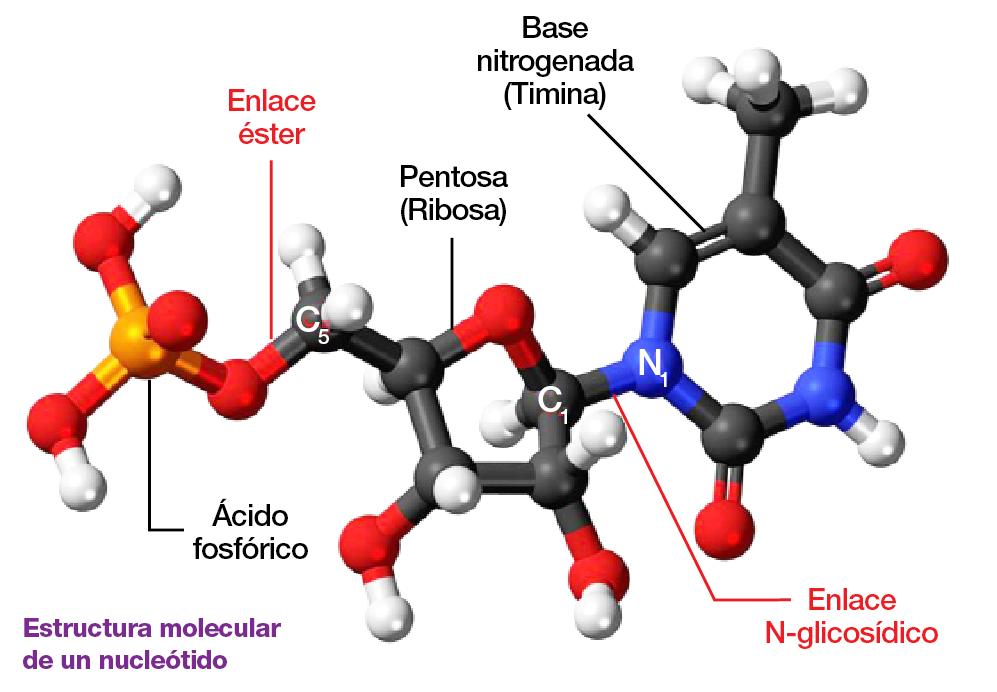
La partición clásica entre los compuestos moleculares de una planta reconoce cuatro bio o macromoléculas básicas: los hidratos de carbono, las proteínas, los lípidos y los ácidos nucleicos, compuestos a su vez por moléculas llamadas nucleótidos. Y es a los nucleótidos a los que en este tramo del viaje nos tocará prestar atención.
El ovillo maravilloso
Sabemos también que esos complejos mecanismos, sujetos a variaciones inducidas por la relación de cada organismo vivo con los estímulos del contexto en el que se desarrolla, posee en el genoma su órgano rector superior: el ADN, el cerebro bioquímico de la célula de todo organismo vivo. Una estructura compuesta por moléculas de ácidos nucleicos, sede de la memoria identitaria de la variedad de la especie, responsable de su transmisión hereditaria y de regular y propiciar la producción de proteínas responsables de las diferentes funciones metabólicas que tendrán lugar en el citoplasma. Funciones regulares o excepcionales de acuerdo a las necesidades de respuesta de la planta ante esos estímulos ambientales, en ese lugar en el que está obligada a cumplir el mandato genético de su ciclo vital.
Estos genes están conformados por una secuencia combinatoria de 4 bases o nucleótidos (Adenina, Timina, Citosina y Guanina), moléculas que codifican de acuerdo a la misión de cada gen, cifrada precisamente en su clave genética, determinada por esa combinación.
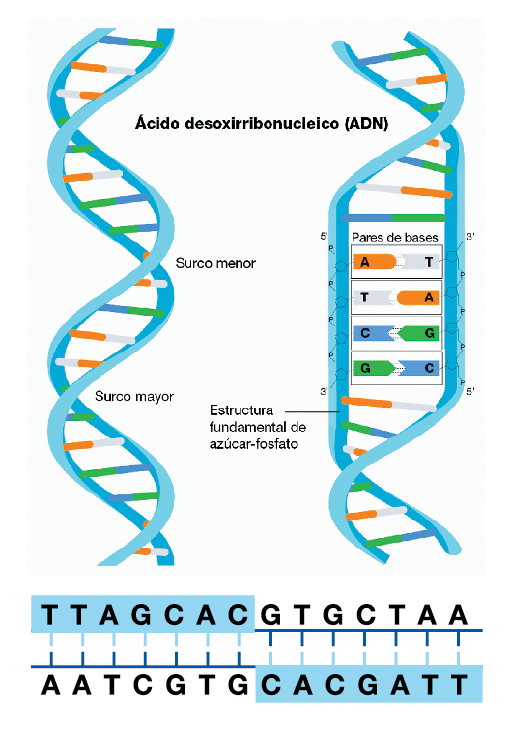
En las células eucariotas estas cadenas de nucleótidos están dispuestas en dos largas hebras en las que cada base de una se combina con su complementaria, siempre en la misma relación de pareja: la Adenina con la Timina (A-T) y la Citosina con la Guanina (C-G). Esta doble cadena se encuentra enrollada sobre sí misma en forma de hélice, y comprimida, de modo que no todos los genes que la componen están expuestos todo el tiempo y en actividad. Si le toca, la región del ADN en la que se encuentre el gen requerido, se desenrollará para exponerlo y expresarlo.
Sabemos también que no todos los genes cumplen funciones similares, que hay regiones genéticas que son codificantes y otras que, en cambio, son promotoras de esa codificación; que en el lenguaje genético operan proteínas intermedias que avisan a los promotores de la necesidad de activar la función codificante de otro gen, proteínas que a su vez han sido codificadas con ese objetivo también por genes que han recibido alguna señal del citoplasma que los pone en alerta. Y sabemos también que en ese ir y venir de señales proteicas intervienen moléculas de otro compuesto genético, los ARN, que cumplen, entre otras, funciones de mensajería o de transcripción: traducen la indicación génica y activan su cumplimiento.
Es muchísimo lo que se ha avanzado en el conocimiento de estos procesos moleculares en las últimas tres décadas, pero también es mucho lo que resta por conocer. Para nuestros fines ahora, diremos que en ese alfabeto limitado de cuatro letras (A, T, C, G) se cifran millones de características de los seres vivos y que el cambio, o la supresión de una sola, o toda una “frase” en esas secuencias modificará, para bien o para mal, el comportamiento de un individuo vegetal ante una situación dada. Esas mutaciones ocurren naturalmente: un gen puede silenciarse o activarse según la circunstancia en la que se encuentre la planta. Pero se ha visto que es posible inducirlas voluntariamente mediante la ingeniería genética. Y en eso estamos.
Corte y reparación
La edición genómica opera de dos maneras diferentes. Siempre se tratará de efectuar un corte en la secuencia de nucleótidos de una o las dos hebras del ADN; y siempre será la propia planta la que repare el corte: o uniendo por las suyas los extremos que han quedado (NHEJ: unión de extremos no homólogos), cambiando la secuencia original y causando la inactivación del gen: o utilizando un “molde” que le aportaremos nosotros, para una reparación por homología (HDR) incorporando la secuencia con la que se reemplazará la que habremos eliminado.
En el primer caso, la reparación realizada por la planta ante la emergencia producida por esa supresión dará como resultado la anulación de la función del gen afectado, pero la planta habrá evitado el riesgo de una descomposición general de su desarrollo; en el segundo, si hemos diseñado bien el reemplazo necesario, y definido en reemplazo de qué, lograremos un cambio deseado en la codificación del gen, que se activará dotando a la nueva planta de la aptitud pretendida.

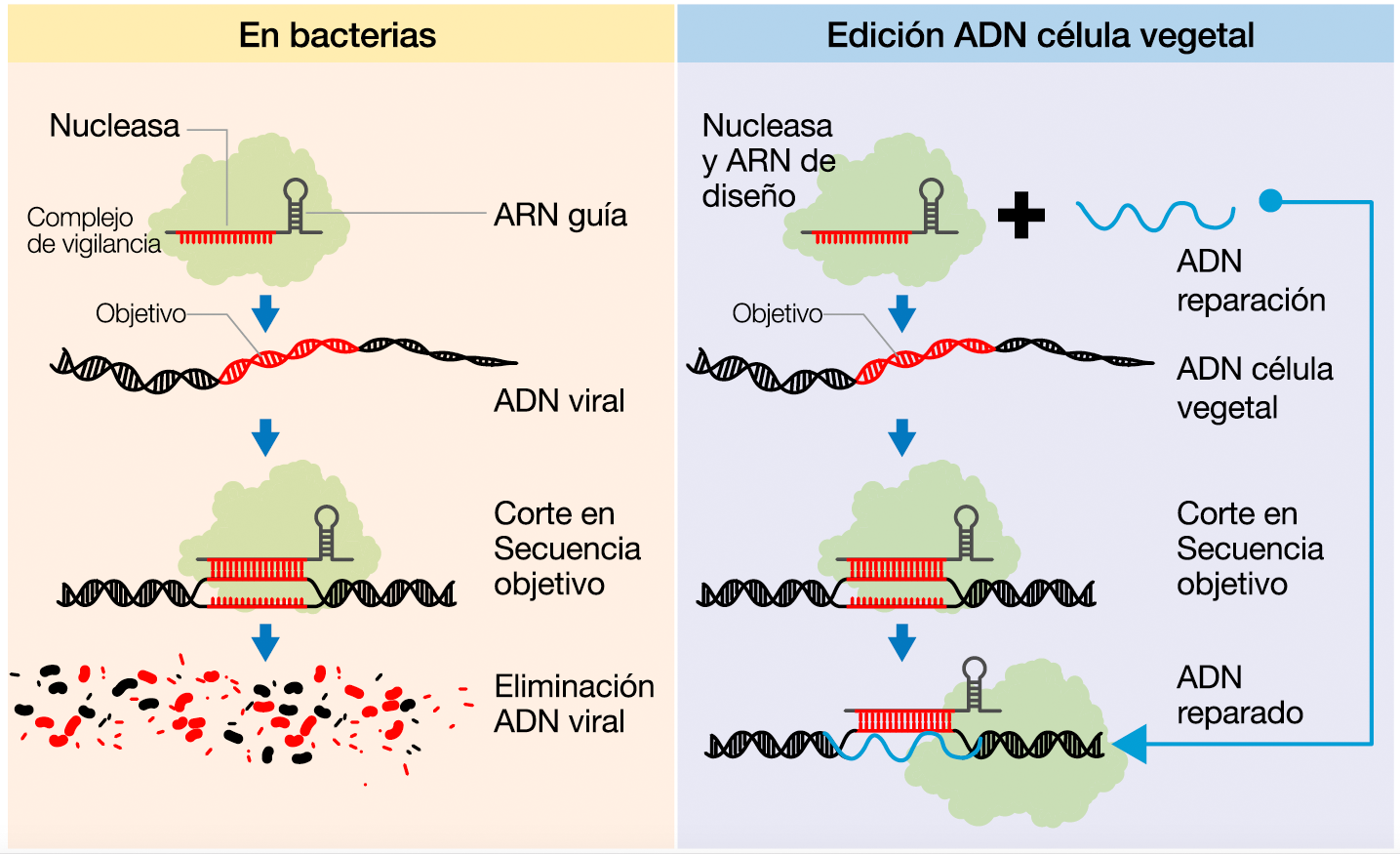
Las herramientas
Participaremos del procedimiento mayormente utilizado en los laboratorios del Itanoa donde nos encontramos: la biobalística, el bombardeo de alta presión de partículas de oro recubiertas por las tres herramientas de diseño aptas para la operatoria. Dos de ellas similares a las que integran el complejo de vigilancia bacteriano conocido como sistema CRISPR-Cas y sirven para efectuar el corte del genoma en el lugar deseado; y una tercera, el ADN de reparación: ese molde de una sola hebra que permitirá a la planta homologar esos extremos, reemplazar la secuencia y mantener, con ese cambio incluido, la actividad del gen en cuestión.
El objetivo
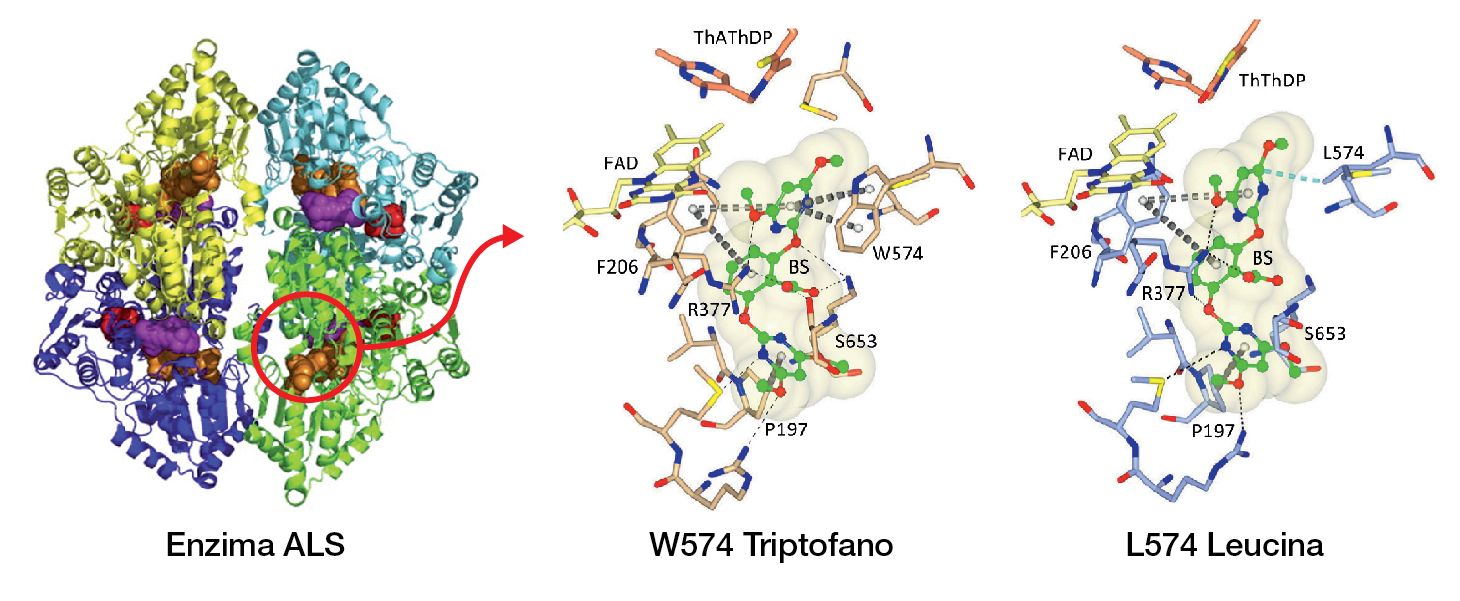
El bombardeo está por comenzar. El objetivo deseado es, como dijimos, modificar una secuencia del ADN de células de caña de azúcar que codifica para una enzima, la ALS -acetolactato sintasa-, participante esencial en el crecimiento de la planta.
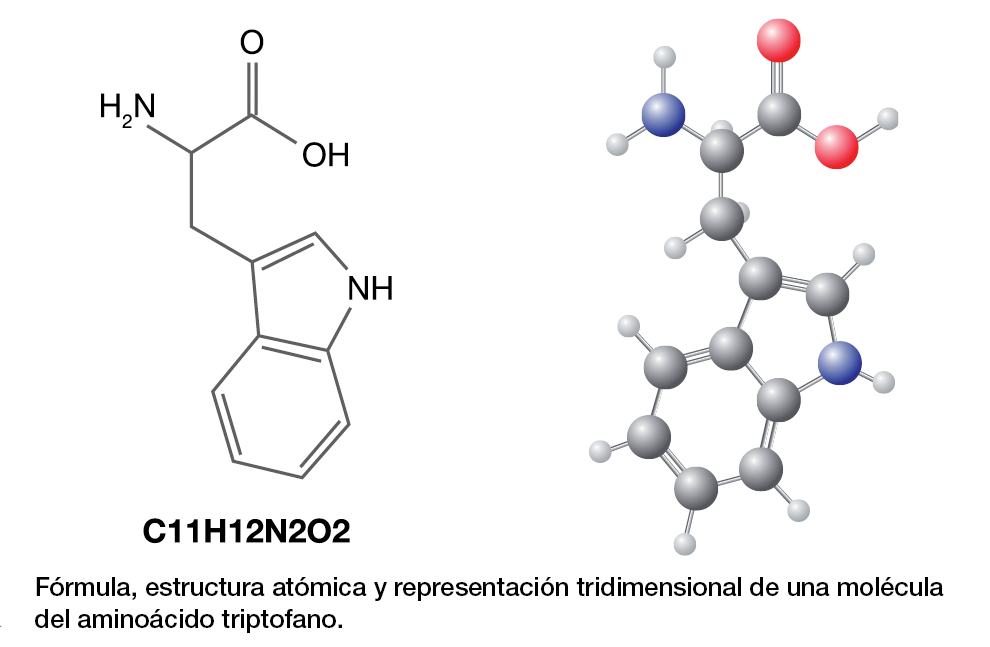
Esa enzima es fuertemente inhibida por herbicidas. Entre las moléculas que integran su cadena polipeptídica participa el aminoácido triptofano; hoy se sabe que deja de ser vulnerable si se elimina ese aminoácido de su composición y se lo sustituye por otro; en este caso, será la leucina. Ese será el objetivo esta vez: lograr obtener una variedad resistente a herbicidas.
Apunten…
Comienza el bombardeo. Miles de proyectiles de oro recubiertos por el material genético diseñado impactan ahora en otras tantas células de caña de azúcar dispuestas en cajas de Petri con el propósito de alcanzar en su núcleo la secuencia de ADN a modificar. Conducidos por el ARN guía, la nucleasa destinada a cortar la secuencia objetivo y el material genético de reemplazo buscarán impactar en la región de ADN “marcada” por otra secuencia adyacente, llamada PAM (Protospacer Adjacent Motifs), que será la que señale al complejo CRISPR-Cas dónde efectuar el corte.
El proceso no está exento de cierto dramatismo. Esos proyectiles con el material genético predispuesto deberán atravesar la membrana celular, el citoplasma, luego la doble membrana nuclear e impactar en el ADN antes de que las proteasas propias de las células, que actúan ante la presencia de material extraño, destruyan el complejo de recambio. No todos los proyectiles darán en el blanco y no todos los que sí lo hagan podrán con el delivery genético buscado.
Lo que sigue
¿Lo habremos logrado esta vez? Solo de unas pocas de esas células exitosamente impactadas podrá prosperar una nueva planta con las características deseadas. Si se identifican esas células promisorias, comenzará con ellas el proceso de fenotipado -de observación, análisis y comprobación efectiva, en laboratorio y en campo- del que resulte una variedad que exprese probadamente la nueva propiedad.
Logrado ese fundamental objetivo y puesta posteriormente a producir y para eso a reproducirse, la aventura de esa nueva planta recién habrá comenzado. Con la leucina en lugar del triptofano en la composición de su enzima ALS, es cierto, para evitar ser dañada por herbicidas de uso todavía necesario, pero expuesta como cualquier otra variedad de la misma especie a las condiciones, bióticas y abióticas, del contexto en el que será obligada a vivir. Una aventura biológica que supone un constante ir y venir de señales intracelulares, en un diálogo permanente entre el citoplasma y el núcleo, con efectos -llamados epigenéticos- en la expresión y la memoria del genoma que, como entendimos, posee su propia capacidad de adaptación.
Al encuentro de dos caminos
A los ojos de las necesidades productivas el tiempo cuenta. Las especies vegetales que se cultivan en gran escala para dar de comer requieren atenciones especiales en un presente continuo. Las plantas irán aprendiendo, es cierto, pero apurarlas un poco se justifica. El desafío es lograr variedades provistas de sus propios recursos para resistir, crecer y brindar sus frutos ahí donde decidimos plantarlas. Hoy entendemos que cuanto más amigables sean nuestros procederes con la naturaleza, mejor; y el conocimiento para conseguirlo crece exponencialmente en nuestros días.
Evolucionamos. A la luz de lo que hoy se sabe que ocurre en el citoplasma durante el crecimiento de una planta en un ambiente determinado, una hipótesis integradora viene cobrando fundamento entre las ciencias que se ocupan de investigar ese universo. La simultaneidad con las que las distintas vías de señalización por las que corren las instrucciones operativas de una punta a la otra de una planta, de acuerdo a la circunstancias ambientales de su desarrollo y a su capacidad heredada , lleva a pensar en la existencia de una matriz, sensible y reactiva, obrando ahí como un tablero inteligente de control, integrador genoma-metabolismo. Una “conciencia superior”, desde la que se dispararían las diferentes vías de señalización que regulan la respuesta de la planta como un todo, en cada momento de su desarrollo.
La hipótesis de una “inteligencia cósmica” que integra las capacidades y las instrucciones del genoma con el desempeño metabólico de la planta, en un flujo constante de señales de ida y vuelta, ayuda a orientar el necesario encuentro de dos ‘tribus’ operantes en el sistema científico actual: la de la ingeniería genética y la de la bioquímica molecular, con sus respectivos subcomponentes: los del lado del núcleo y los del citoplasma” (ellas y ellos, obviamente, en ambos casos). Un encuentro que permita definir los retoques genéticos que quepan en función de lo que integralmente significarán para la planta como un todo, dotándola de la mayor capacidad posible para resistir en su lugar, para aprender y reaccionar positivamente también ante el estímulo circunstancial de bioinsumos moleculares que, cual vacunas, refuercen su memoria y su aptitud natural.
Al decir de los que de esto saben, no estaríamos tan lejos de lograrlo: ¿10, 15 años? Si esa alianza de miradas encuentra su punto de encuentro “núcleo” y excava en consecuencia como es esperable, ese horizonte sería alcanzable y ese lapso, posible.
Rebote
Hemos llegado al núcleo, fin del camino de ida. Toca emprender el de regreso. De salida, tendremos que atravesar de nuevo el citoplasma. El tránsito por ahí es intenso y suele congestionarse. Tendremos que afinar los parámetros de nuestro GPS para ver por dónde ir.
![]()